Itsas ugaztun
Itsas ugaztunak ozeano- eta itsas ekosistemetan bizi diren ugaztun urtarrak dira. Talde honen barruan sartzen dira itsas txakurrak, baleak, manatiak, itsas igarabak eta hartz zuriak. Itsas ingurunean elikatzeko beharra da animalia horiek talde bakarrean elkartzen dituen ezaugarri bakarra.


Itsas ugaztunen kopurua txikia da lehorrean bizi diren ugaztunekin konparatuta. Itsas ekosistemetan dituzten zereginak askorarikoak dira, besteak beste, itsas ekosistemen mantentzean eta harrapakinen erregulazio-prozesuetan hartzen dute parte.
Aborigeneak izan ziren lehen aldiz itsas ugaztunak ehizatu zituzten pertsonak, haietatik elikagaiak eta beste baliabide asko lortzeko. Beste itsas ugaztun asko industriaren eta merkataritzaren oinarria izan ziren, eta ustiatutako espezieen populazioen gainbehera azkarra gertatu zen, batez ere zigilu eta baleena. Harrapakaritza komertzialak hainbat espezieren iraungipenera eragin du, adibidez, itsas bisoiena, Japoniako itsas lehoiena eta Karibeko itsas txakur mediterranearrena. Harrapaketa komertziala amaitu zenean, hainbat espezieren kopurua handitu egin zen, iparraldeko elefante zigiluarena eta bale grisarena besteak beste. Egun, beste animalia batzuek arrisku larrian jarraitzen dute, Ipar Atlantitkoko baleak adibidez. Dena den, nahiz eta harrapakaritza komertziala amaitu, itsas ugaztun asko hil daitezke nahigabe arrantzarako erabiltzen diren sareetan harrapatuta geratzen direnean. Ozeanoetako trafikoa handitzeak itsasontzien eta ugaztun handien arteko talka maizago gertatzea eragin du. Habitataren degradazioa animalia hauek jasaten duten beste mehatxu bat da, elikagaiak bilatzeko eta harrapatzeko gaitasunean eragiten baitu. Kutsadura akustikoak, adibidez, animalien ekolokalizazioan eragin dezake, eta berotze globalaren eraginak artikoetako inguruneak suntsi ditzake.
Taxonomia
aldatu



.jpg)
Existitzen diren espezieen sailkapena
aldatu- Cetartiodactyla ordena[1]
- Whippomorpha subordena
- Balaenidae familia (baleak), bi genero eta lau espezie
- Cetotheriidae familia (balea), espezie bakarra
- Balaenopteridae familia (baleak), bi genero eta zortzi espezie
- Eschrichtiidae familia (bale grisa), espezie bakarra
- Physeteridae familia (kaxalotea), espezie bakarra
- Kogiidae familia (baleak), genero bakarra eta bi espezie
- Monodontidae familia (narbala eta beluga), bi genero eta bi espezie
- Ziphiidae familia (baleak), sei genero eta hogeita bat espezie
- Delphinidae familia (itsas izurdeak), hamazazpi genero eta hogeita hemezortzi espezie
- Phocoenidae familia (mazopak), bi genero eta zazpi espezie
- Whippomorpha subordena
- Sirenia ordena (itsasoko behiak)[1]
- Trichechidae familia (manatiak), bi espezie
- Dugongidae familia (itsasuskia), espezie bakarra
- Carnivora ordena (haragijaleak)[1]
- Caniformia subordena
- Mustelidae familia, hiru espezie
- Ursidae familia (hartza), espezie bakarra
- Pinnipedia subordena (itsas lehoiak, mortsak, itsas txakurrak)
- Otariidae familia (belarridun itsas txakurrak), zazpi genero eta hamabost espezie
- Odobenidae familia (mortsa), espezie bakarra
- Phocidae familia (belarri-gabeko itsas txakurrak), hamalau genero eta hemezortzi espezie
- Caniformia subordena
Eboluzioa
aldatu
Itsas ugaztunak 129 espeziek osatzen duten askotariko taldea da, eta bizi ahal izateko ozeanoa behar dute[2][3]. Talde horiek soilik biltzen dira talde batean itsas ingurunea elikatzeko behar dutelako eta horrekiko mendekotasuna dutelako[4]. Itsas inguruneen beharrizan maila asko aldatzen da espeziearen arabera. Itsas ugaztunek ur-ingurunean bizitzeko garatzen dituzten moldapenak oso desberdinak dira baita espezieen arabera ere[5][6]. Zetazeoak eta sirenioak guztiz urtarrak dira eta, ondorioz, uretako bizimodua izatera behartuta daude. Baleak eta itsas lehoiak, aldiz, erdiurtarrak dira eta, haien denbora gehiena uretan ematen duten arren, lehorrera bueltatu behar dira estalketa, muda edo haziera moduko funtzio garrantzitsuak betetzeko. Azkenik, itsas igarabek eta hartz zuriek ez dute ur-ingurunean bizitzeko horrenbeste moldapen. Mota desberdinetako itsas ugaztunen dieta ere askotarikoa da. Batzuk zooplanktonaz elikatzen dira, beste batzuk arrainez, txipiroiz, itsaskiz eta algez, eta gutxi batzuk bestelako ugaztunez elikatzen dira[4].
Zetazeoak urtar bilakatu ziren duela 50 milioi urte inguru[7]. Ikerketa molekularretan eta morfologikoetan oinarrituta, zetazeoak genetikoki eta morfologikoki Artiodactyla, behatz kopuru bikoitia duten ungulatuak, ordenaren barruan kokatu dira[8][9]. "Cetartiodactyla" hitzak esan nahi du baleak ungulatuetatik eratorri zirela. Izena bi ordenak, Cetacea eta Artiodactyla, elkartzetik dator. Definizio horren barruan, baleen eta izurdeen ahaide lehortar gertukoena hipopotamoa dela uste dute ikertzaileek[10][11][11][12].
Sirenioak, itsasoko behiak, itsastar bilakatu ziren duela 40 milioi urte inguru. Sirenioen lehen agerpena fosilen erregistroan Eozeno goiztiarrean izan zen, eta Eozeno bukaerarako esanguratsuki dibertsifikatu ziren. Ibaietako, estuarioetako eta itsas kostako animalia urtarrak azkar zabaltzeko gai izan ziren. Beste itsas ugaztun batzuk ez bezala, Sirenio zaharrena, †Prorastomus, Jamaikan aurkitu zen[6]. Ezaguna den Eozeno goiztiarreko lehen Sirenio tetrapodoa †Pezosiren izan zen[13]. Ezagunak diren lehen itsas behiak, Prorastimidae eta Protosirenidae familiatakoak, Eozeno garaikoak dira, eta txerri tamaina zuten lau hankako anfibioak ziren[14]. Dugongidae familiako lehen kideak Eozeno garaiaren bukaeran agertu ziren[15]. Orduan, itsas behiak guztiz urtarrak ziren jada[14].
Eozeno garaian, pinipedioak beste kaniformeetatik banandu ziren duela 50 milioi urte. Ugaztun lehortarrekin duten lotura ebolutiboa ezezaguna zen 2007ra arte non Miozeno goiztiarreko †Puijila darwini-ren gorotzak aurkitu ziren Nunavut-en (Kanada). Igaraba modernoek bezala, Pujiila-k isats luzea, gorputz-adar motzak, eta hegatsen ordez sare moduko oinak zituen[16]. Otariidae (belarridun itsas txakurrak) eta Odobenidae (mortsak) leinuak duela 28 milioi urte banandu ziren[17]. Fozidoak (belarri gabeko itsas txakurrak) duela 15 milioi urtetatik existitzen dira gutxienez, eta ebidentzia molekularrek erakusten dute Monachinae (itsas txakur fraileak) eta Phocinae leinuak duela 22 milioi urte banandu zirela[17].
Ebidentzia fosilek erakusten dute itsas igaraben (Enhydra) leinua Ipar Pazifikoan isolatu zela duela 2 milioi urte gutxi gorabehera, orain iraungitako Enhydra macrodonta eta itsas igaraba modernoa (Enhydra lutris) emanda. Hasieran, itsas igarabek ipar Hokkaido eta Errusian eboluzionatu zuten, eta geroago zabaldu ziren ekialdera, irla Aleutianetara, Alaskara, eta Ipar Amerikako kostatik behera. Itsas igaraba nahiko berria da itsasoko bizitzan duela 50 milioi urte sartu ziren zetazeoekin, duela 40 milioi urte sartu ziren sirenioekin eta duela 20 milioi urte sartu ziren pinipedoekin alderatuta. Alderdi batzuetan, ordea, itsas igaraba hobeto egokituta dago itsas bizitzara pinipedoak baino, pinipedoak lehorrera edo izotzetara joan behar baitira erditzera[18].
Hartz zuriak hartz arreetatik (Ursus arctos) eratorriak direla uste dute ikertzaileek: Pleistozeno[19] garaiko glaziazio batean hartz arrea isolatu ondoren hartz zuri bilakatu zela. Ezaguna den hartz polar fosil zaharrena duela 130.000-110.000 urteko baraila-hezurra da, Prince Charles Foreland-en aurkitua 2004an[20]. Hartz polarraren mitokondrio-DNA (mtDNA) hartz arrearenetik bereizi zen duela 150.000 urte[20]. Gainera, hartz arre talde batzuek erlazio estuagoa dute hartz zuriekin beste hartz arre batzuekin baino[21], beraien mtDNAren arabera; horrek esan nahi du hartz zuriak agian ez liratekeela espezietzat hartu behar[22].
Oro har, lehorreko amniotoen itsasoaren kolonizazioa Zenozoikoan maizago gertatu zen Mesozoikoan baino. Joera hori bultzatu zuen faktorea izan zen kostatik gertu zeuden itsas inguruneen emankortasunaren handipena. Trantsizioa erraztu zuen endotermia ere erabakigarria izan zen[23].
Banaketa eta habitata
aldatu
Itsas ugaztunak modu zabalean daude banatuta lur planetan zehar, nahiz eta haien banaketa ozeanoen produkzioarekin bat egiten duen[24]. Espezieen aberastasun handian 40º-ko latitudeen inguruan ageri da, bai iparraldean eta bai hegoaldean. Ipar eta Hego Amerika, Afrika, Asia eta Australia inguruan agertzen den ekoizpen primario handiaren ondorioz gertatzen da. Espezieen hein totala aldakorra da itsas ugaztun espezieen arabera. Batez beste, itsas ugaztunek Ozeano Indikoaren azaleraren 1/5a betetzen dute.[25] Ikusitako tamainaren aldakortasuna baldintza ekologiko desberdinen eta ingurumen-baldintza aldakorrei aurre egiteko duten gaitasunaren araberakoa da. Itsas ugaztunen espezieen aberastasun handiaren eta gizakiak ingurunean duen eraginaren arteko harremanak gainjarri egiten dira[2].
Itsas ugaztun gehienak, itsas txakurrak eta itsas igarabak besteak beste, kostaldean bizi dira. Itsas txakurrek habitat lehortarrak ere erabil ditzakete, bai uharteetan eta bai ingurune kontinentalean. Ingurune tropikaletan, hondartza hareatsu eta harritsuetan, itsas kobazuloetan, mareek sortutako putzuetan edo hondar-bankuetan biltzen dira ugalketarako edo atseden hartzeko. Beste espezie batzuek gizakiak sortutako plataformetan ere hartzen dute atseden[26]. Itsas txakurrak ingurune lehortarrean mugitu daitezke, hareazko dunetan atseden hartu eta itsaslabarrak igo ahal dituzte[27]. Zetazeo gehienak, bestalde, itsaso zabalean bizi dira, eta kaxalote moduko espezieak 300-760 m bitarteko sakoneretan igeri egin dezakete janaria bilatzeko. Sirenioak kostaldean bizi dira, sakonera txikiko uretan, 9 m-ra itsas mailaren azpian[28]. Dena den, Sirenio batzuk sakonera handiagoetan algak bilatzen ikusi dira. Itsas igarabak babestutako eremuetan bizi dira, hesi-arrezifeetan edo kostalde arrokatsuetan[29]. Dena den, izotz mugikorretan edo gune areatsuetan ere egon daitezke[30].
Itsas ugaztun askok migratu egiten dute urtaroen arabera. Baleek eguraldi-aldaketen ondorioz agertu eta desagertu egiten diren izotz blokeen arabera migratzen dute. Gainera, balea batzuek distantzia luzeko migrazioak egiten dituzte, ur tropikaletan erditu eta kumea hezi ahal izateko. Hartz zuriek, aldiz, haien harrapakinei jarraituz migratzen dute. Badia batzuetan, urteko aldi batzuetan izotza guztiz desagertzen da eta hartz zuriek bere burua behartua ikusten dute lehorrera migratzeko, hurrengo izozte-garaia heltzen den arte[31].
Moldapenak
aldatu
Itsas ugaztunek bizitza urtarreko hainbat ezaugarri fisiologiko eta anatomiko gainditu behar dituzte. Ezaugarri horietako batzuk berezkoak dira espezie batean. Itsas ugaztunek hainbat ezaugarri garatu dituzte lokomozio eraginkorra lortzeko, hala nola, arrastea ekiditeko torpedo formako gorputzak, propultsiorako eta gidatzeko gorputz-adar eraldatuak, eta propultsiorako eta orekarako isats- eta bizkar-hegatsak[25]. Itsas ugaztunak termorregulazioan adituak dira, ilaje dentsoak, gantzak, edo odol-zirkulazioaren doikuntzak baitituzte; gainera, torpedo formako gorputzek, gorputz-adar motzek, eta tamaina handiak bero-galera ekiditen laguntzen diete[25].
Itsas ugaztunak urpean denbora luzez egoteko gai dira. Pinipedioek eta zetazeoek odol-hodien sistema handiak eta konplexuak dituzte, urperatze sakonak egiteko beharrezko oxigenoa metatzeko. Beste metaketa-gune batzuk muskuluak, odola, eta barea dira, oxigeno-kontzentrazio handiak metatzeko gaitasuna dutenak. Bradikardia, hau da, bihotz-taupaden erritmoa moteltzea, eta basokonstrikzioa, oxigeno gehiena garuna eta bihotza bezalako berebiziko organoetara bideratzea, egiteko gai dira, denbora luzeagoz urperatuta mantentzeko eta oxigeno falta gainditzeko[25]. Oxigenoa agortzen bada, itsas ugaztunek glukogeno-erreserba ugari dituzte, murgiltze luzeetan gertatzen diren hipoxia-egoeretan glukolisi anaerobikoa egiteko[32][33].
Soinuak desberdin bidaiatzen du uretan; horregatik, itsas ugaztunek hainbat egokitzapen garatu dituzte komunikazio, ehiza, eta harraparien detekzio eraginkorrak lortzeko[34]. Baleen eta izurdeen egokitzapen esanguratsuena ekolokalizazioaren garapena da[25]. Horzdun baleek maiztasun altuko izpi-pultsuak igortzen dituzte beraien burua begira dagoen noranzkoan. Soinuak hezurrezko sudur-hobietako airea ezpain fonikoetatik bideratuz eratzen dituzte[35]. Soinu horiek garezurreko hezur konkabo dentsoak eta haren oinarrian dagoen aire-zaku batek islatzen dituzte. Izpi enfokatu hori “meloi” izeneko organo gantzatsu batek modulatzen du. Organo gantztsuak leiar akustiko bezala jokatzen du, dentsitate desberdinetako lipidoz osatuta baitago[35][36].
Itsas ugaztunek ezaugarri ugari garatu dituzte elikatzeko, beraien hortzaldian ikus daitezkeenak. Esaterako, pinipedioen eta odontozetoen masail-haginak propio moldatuta daude arrainak eta txibiak harrapatzeko. Alderantziz, bale-bizarrak planktona eta arrain txikiak iragazteko garatu dira[25].
Hartz zuriek, itsas igarabek, eta itsas txakur iletsuek ilaje luzea, oliotsua, eta iragazgaitza dute airea harrapatu eta isolamendu termikoa ahalbidetzeko. Baleek, izurdeek, mazopek, manatiek, itsasuskiek, eta mortsek, ordea, ilaje luzea galdu dute, eta epidermis lodi eta dentsoa eta gantz-geruza lodia garatu dute, arrastea ekiditeko. Itsas hondoan elikatzen direnak, manatiak adibidez, ura baino pisutsuagoak izan behar dira hondoarekin kontaktua mantentzeko edo urperatuta egoteko. Itsas azalean bizi diren animaliek, itsas igarabek adibidez, alderantzizkoa behar dute, eta animalia igerilariek, izurdeek esaterako, flotagarritasun neutroa behar dute uretan gora eta behera ibiltzeko. Oro har, hezur lodi eta dentsoak dituzte itsas hondoko animaliek, eta dentsitate baxuko hezurrak, aldiz, ur sakonetan bizi diren ugaztunek. Hartz zuriak eta itsas igarabak bezalako itsas ugaztun batzuk pisua mantentzen duten gorputz-adarrak mantendu dituzte, eta horiei esker lurrean ibiltzeko gai dira, lehorreko animaliek bezala[37].
Ekologia
aldatuDieta
aldatuZetazeo guztiak dira harrapakariak eta haragijaleak. Haginak dituzten baleak arrainez eta zefalopodoz, gehienbat, eta krustazeoz eta bibalbioz elikatzen dira. Beste balea-espezie batzuk eta ponipedio-espezie batzuk iragazleak dira[26][38]. Partekatzen duten harraparitza-metodo bat arrainzaintza da, zeinean arrain-bolumen txiki batez arduratzen diren. Ondoren, taldeko kide bakoitzak arrantza egiten du, zaindutako arrainez elikatzeko[39]. Izurdeek erabiltzen duten arrantza-mekanismoa desberdina da, azaleko uretan dauden arrainez elikatzen dira, harrapatzeko errazagoak direlako[39]. Orka arruntek eta izurde handiek haien harrapakina kostaldera bideratzen dute bertan ehizatu ahal izateko. Orka arruntak ezagunak dira marrazo zuriak eta arraiak geldiarazteko gai direlako. Haragijaleak diren arren, haien hesteetako mikrobiota herbiboro lehortarren antzekoa da, seguruenik aitzindari belarjale bat dutelako[40].

Balea bizardunek beren bizar-plaka erabiltzen dute planktona harrapatzeko; bi metodo erabiltzen dituzte horretarako: lunge-elikadura eta trago-elikadura. Lunge-elikadura egiten duten animaliek beren baraila luzatzen dute ahoa emendatu ahal izateko. Luzatze horrek haien eztarrian ildoak eratzen dituelarik, barneratu dezaketen ur-bolumena handitu egiten da[41][42]. Metodo hori bakarrik izaten da energetikoki eraginkorra elikagai-masa handi baten aurrean aplikatzen dutenean. Trago-elikadura egiten duten animaliek ahoa irekita egiten dute igeri eta, horrela, ura eta harrapakinak barneratzen joaten dira. Kasu honetan, harrapakinek tamaina egokia eta igeriketa geldoa izan beharko dute harrapatuta geratzeko.
Igarabak dira arroken artean mugitzeko gaitasuna duten itsas animalia bakarrak. Igarabek barraskiloen moduko organismoak jasotzen dituzte hondoko urtean bibalbioen bila murgildu ahal izateko. Atzaparrekin ehizatzen duen itsas ugaztun bakarrak dira: gorputz-adar bakoitzaren azpian poltsa bat dute elikagaiak bildu ahal izateko eta azalera garraiatu ahal izateko[43]. Igarabek beren soinaren gainean flotatzen duten bitartean elikatzen dira, poltsetan gorde duten elikagaiak atzaparrekin hartuz eta ahora eramanez[44]. Oro har krustazeoez eta arrainez elikatzen dira[45].
Pinapidoak krustazeoz, bibalbioz, zefalopodoz arrainez, zooplanktonez eta odol-beroko harrapakinez elikatzen dira[26] eta oro har, ez dira oso espezifikoak[46]. Normalean ornogabe sesilez, taldeetan antolatzen diren animalia endodermikoez edo animalia geldoez elikatzen dira. Bakartiak diren espezieak kostaldeko uretan, badietan edo erreketan aurkitzen dira. Arrain-banku handiak aurkitzen dituztenean, modu kooperatiboan ehizatzen dute[26].

Hartz zuria hartz-espezie karniboroena da. Oro har itsas txakur marmolairez eta itsas txakur bizardunez elikatzen dira hartz zuriak[47]. Izotzaren, uraren eta airearen arteko interfasean ehizatzen dute batez ere; zigiluak lehorrean edo uretan ehizatzen dituzte bakarrik[48]. Ehizatzeko, usaimenaren bidez zigiluen arnasketarako zuloak hautematen dituzte eta horien inguruan makurtzen dira zein ateratzen diren arte. Hartz zuriak zigiluaren arnasketa nabaritzen duenean, bere atzaparra zuloan sartu egiten du, zigilua gordelekutik ateratzeko. Hartz zuriek ehizatzeko erabiltzen duten beste mekanismo bat da izotzean etzanda dauden zigiluak ehizatzea[49]. Erabiltzen duten azken mekanismoa, emeek erditzeko elurretan eraikitzen dituzten gordelekuak erasotzea da[50].
Sirenidoak itsasoko landareez elikatzen dira gehienbat. Landare osoa ingeritzen dute normalean, sustraiak barne. Hori egitea posible ez bada, hostoez elikatzen dira bakarrik[51]. Itsasoko landare mota asko aurkitu dira dugongoen urdailean eta ondorioztatu da, itsasoko belar gutxi daudenean algez elikatzen direla[52].
Funtsezko espezieak
aldatuIgarabak oso espezie garrantzitsuak dira; ekosistemetan duten eragina beraien tamainak eta kopuruak diruditena baino handiagoa da. Belarjale bentiko batzuen populazioak kontrolpean mantentzen dituzte, bereziki itsas trikuen populazioa. Itsas trikuak alga arreez elikatzen dira, eta horien heriotza eragiten dute. Alga arreen basoek eskaintzen duten habitata eta bertako nutrienteak galtzeak ondorio nabariak ditu itsas ekosistemetan. Ipar Pazifikoko gune batzuetan ez dago igarabarik, eta ondorioz, itsas trikuen ekosistema ez-emankorrak agertzen dira, itsas triku ugarirekin eta alga arreen basorik gabe[53]. British Columbian igarabak berriz sartzeak nabariki hobetu du itsas ekosistemen osasuna[54]. Aleutiarretan, Komandante uhartetan, eta Kaliforniako kostan igarabak berriz sartuz antzeko emaitzak lortu dira. Hala ere, Kaliforniako alga arreen baso batzuk berreskuratu egin dira igarabak sartu gabe; beraz, itsas trikuen populazioa beste faktore batzuek kontrolatzen dute gune horietan[55]. Igarabek itsas trikuen populazioak kontrolatzeko duten gaitasuna garrantzitsuagoa da kosta irekietan babestuago dauden badia eta estuarioetan baino[55].
Goi-harrapakariek eragina dute harrapakinen populazio-dinamikan eta defentsa-estrategietan (kamuflajean adibidez)[56]. Hartz zuria da goi-harrapakari nagusia[57]. Beste hainbat espeziek, bereziki azeri artikoek (Vulpes lagopus) eta ipar-kaioek (Laur hyperboreus), hartz zuriek hildako hondarrak jaten dituzte[58]. Hartz zurien eta itsas txakur marmolaireen arteko harremana hain estua da, ezen itsas txakur marmolaireen ugaritasunak hartz zurien dentsitatea erregulatzen duen, eta era berean, hartz zurien harrapakaritzak itsas txakur marmolaireen dentsitatea eta ugaltze-arrakasta erregulatzen dituen[48]. Hartz zuriek itsas txakurrez elikatzeak eragindako presio ebolutiboak itsas txakur artikoen eta antartikoen arteko desberdintasunak azaltzen ditu[58]. Ur-azaleko harrapakaririk ez duten itsas txakur antartikoek ez bezala, itsas txakur artikoek arnas-zulo gehiago dituzte, atseden gutxiago hartzen dute izotz gainean dauden bitartean, eta oso gutxitan sabelusten dute izotz gainean. Itsas txakurkume artikoen ilajea zuria da, seguraski kamuflajerako, itsas txakurkume antartikoen ilajea, aldiz, iluna da[58].
Orkak goi-harrapakariak dira bizi diren gune guztietan, eta eragin nabariak izan ditzakete harrapakinen espezieetan. Beraien dieta oso zabala da, eta hainbat itsas ornodunez elikatzen dira: izokinak[59], arraiak, marrazoak (baita marrazo zuriak ere)[60][61], balea handiak[62], eta 20 pinipedo espezie barne[63]. Orkak balea jaioberriez elikatzen dira, eta hori baleak ur tropikaletara migratzearen eta bertan kumeak erditzearen eragilea izan daiteke, ur tropikaletako orka kopurua ur polarretakoa baino askoz baxuagoa delako. Gizakien baleen ehiza baino lehenago, pentsatzen da balea handiak zirela orken elikagai nagusia, baina baleen populazioaren jaitsiera nabariaren ondorioz, orken dieta zabaldu egin zen itsas ugaztun txikiagoetara[64]. 1990eko hamarkadako Aleutiarretako igaraben populazioaren jaitsiera orken harrapakaritzaren ondorio izan zela uste da, baina ez dago hori erakusten duen ebidentzia zuzenik. Itsas igaraben jaitsieraren ondoren, itsas txakur arrunten eta itsas lehoien populazioak jaitsi ziren. Itsas txakur arruntek eta itsas lehoiek jatorrizko harrapakina ordezkatu zuten[65][66][67].
Baleen zikloa
aldatu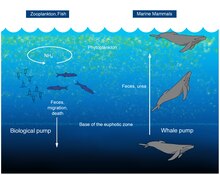
2010eko ikerketa batek erakusten du baleak eragin positiboa dutela arrantzaleengan, “baleen zikloa” izenekoa dela eta. Baleek nitrogenoa bezalako mantenugaiak garraiatzen dituzte itsas hondoetatik azalerara. Garraio horrek goranzko ponpa biologiko bezala jokatzen du. Maineko golkora modu honetan heltzen den nitrogeno kantitatea golkora isurtzen duten ibai guztiek dakarten nitrogeno kantitatea baino handiagoa da, zehazki, 23.000 tona ingurukoa urtero[68]. Baleek ozeanoaren azalean sabelusten dute. Beraien gorotzak garrantzitsuak dira arrantzarako, burdinan eta nitrogenoan aberatsak direlako. Baleen gorozkiak likidoak dira, eta ondoratu beharrean, gainazalean geratzen dira, eta fitoplanktona horietaz elikatzen da[68][69].
Hiltzerakoan, baleen gorpua itsas hondora doa, horrela itsas bizitzarentzat habitat garrantzitsu bat eskainita. Gaur egun ondoratutako balea-gorpuek eta ebidentzia fosilek erakusten dute ondoratutako balea-gorpuek bizidun talde ugari agertzea eragiten dutela, 407 espezietara helduta[70]. Balea-gorpuen degradazioa hiru fasetan gertatzen da. Lehenengo fasean, organismo mugikorrak, marrazoak eta mixinak adibidez, ehun bigunak jaten dituzte hilabete batzuetatik bi urterako denbora-tartean. Ondoren, bigarren fasean, krustazeoak eta poliketoak bezalako oportunista aberasleek hezurrak eta sedimentuak, materia organikoa dutenak, kolonizatzen dituzte urte batzuetan zehar. Azkenik, hirugarren fasean, sufrearen bakterioek hezurrak degradatu, eta hidrogeno sulfuroa (H2S) askatzen dute eta organismo kimioautotrofoen hazkundea ahalbidetzen dute. Organismo kimioautotrofoek aldi berean muskuiluen, txirlen, lapen, eta itsas barraskiloen agerpena eragiten duten. Azken fase honek hamarkadak iraun ditzake, eta batez beste, 185 espezieko gune aberatsak sortzen ditu[71].
Erreferentziak
aldatu- ↑ a b c Taylor, Barbara L.; Perrin, William F.; Reeves, Randall R.; Rosel, Patricia E.; Wang, John Y.; Cipriano, Frank; Scott Baker, C.; Brownell, Robert L.. (2017-06). «Why we should develop guidelines and quantitative standards for using genetic data to delimit subspecies for data-poor organisms like cetaceans» Marine Mammal Science 33 (S1): 12–26. doi:. ISSN 0824-0469. (Noiz kontsultatua: 2019-11-29).
- ↑ a b (Ingelesez) Kaschner, Kristin; Tittensor, Derek P.; Ready, Jonathan; Gerrodette, Tim; Worm, Boris. (2011-05-23). «Current and Future Patterns of Global Marine Mammal Biodiversity» PLOS ONE 6 (5): e19653. doi:. ISSN 1932-6203. PMID 21625431. PMC PMC3100303. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Pompa, Sandra; Ehrlich, Paul R.; Ceballos, Gerardo. (2011-08-16). «Global distribution and conservation of marine mammals» Proceedings of the National Academy of Sciences 108 (33): 13600–13605. doi:. ISSN 0027-8424. PMID 21808012. PMC PMC3158205. (Noiz kontsultatua: 2019-12-12).
- ↑ a b Jefferson, Thomas A.. (2008). Marine mammals of the world : a comprehensive guide to their identification. (1st ed. argitaraldia) Academic ISBN 0-08-055784-8. PMC 326418543. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Uhen, Mark D.. (2007-06). «Evolution of marine mammals: Back to the sea after 300 million years» The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology 290 (6): 514–522. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ a b (Ingelesez) Savage, R. J. G.; Domning, Daryl P.; Thewissen, J. G. M.. (1994-09-07). «Fossil sirenia of the west Atlantic and Caribbean region. V. The most primitive known sirenian, Prorastomus sirenoides Owen, 1855» Journal of Vertebrate Paleontology 14 (3): 427–449. doi:. ISSN 0272-4634. (Noiz kontsultatua: 2019-12-12).
- ↑ Castro, Peter.. (2008). Marine biology. (7th ed. argitaraldia) McGraw-Hill ISBN 978-0-07-302819-4. PMC 155128723. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Geisler, Jonathan H.; Uhen, Mark D.. (2005-06). «Phylogenetic Relationships of Extinct Cetartiodactyls: Results of Simultaneous Analyses of Molecular, Morphological, and Stratigraphic Data» Journal of Mammalian Evolution 12 (1-2): 145–160. doi:. ISSN 1064-7554. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) «Molecular evidence for the inclusion of cetaceans within the order Artiodactyla.» Molecular Biology and Evolution 1994-05 doi:. ISSN 1537-1719. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Agnarsson, Ingi; May-Collado, Laura J.. (2008-09). «The phylogeny of Cetartiodactyla: The importance of dense taxon sampling, missing data, and the remarkable promise of cytochrome b to provide reliable species-level phylogenies» Molecular Phylogenetics and Evolution 48 (3): 964–985. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ a b (Ingelesez) Price, Samantha A.; Bininda-Emonds, Olaf R. P.; Gittleman, John L.. (2005-08). «A complete phylogeny of the whales, dolphins and even-toed hoofed mammals (Cetartiodactyla)» Biological Reviews 80 (03): 445. doi:. ISSN 1464-7931. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Spaulding, Michelle; O'Leary, Maureen A.; Gatesy, John. (2009-09-23). Farke, Andrew Allen ed. «Relationships of Cetacea (Artiodactyla) Among Mammals: Increased Taxon Sampling Alters Interpretations of Key Fossils and Character Evolution» PLoS ONE 4 (9): e7062. doi:. ISSN 1932-6203. PMID 19774069. PMC PMC2740860. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Domning, D. P.. (2001-10). «The earliest known fully quadrupedal sirenian» Nature 413 (6856): 625–627. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2019-12-12).
- ↑ a b Invasion biology and ecological theory : insights from a continent in transformation. ISBN 978-1-107-03581-2. PMC 850909221. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Samonds, Karen E.; Zalmout, Iyad S.; Irwin, Mitchell T.; Krause, David W.; Rogers, Raymond R.; Raharivony, Lydia L.. (2009-12-12). «Eotheroides lambondrano , new middle Eocene seacow (Mammalia, Sirenia) from the Mahajanga Basin, northwestern Madagascar» Journal of Vertebrate Paleontology 29 (4): 1233–1243. doi:. ISSN 0272-4634. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Rybczynski, Natalia; Dawson, Mary R.; Tedford, Richard H.. (2009-04). «A semi-aquatic Arctic mammalian carnivore from the Miocene epoch and origin of Pinnipedia» Nature 458 (7241): 1021–1024. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2019-12-12).
- ↑ a b (Ingelesez) Arnason, Ulfur; Gullberg, Anette; Janke, Axel; Kullberg, Morgan; Lehman, Niles; Petrov, Evgeny A.; Väinölä, Risto. (2006-11). «Pinniped phylogeny and a new hypothesis for their origin and dispersal» Molecular Phylogenetics and Evolution 41 (2): 345–354. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ Love, John A.. (1992). Sea otters. Fulcrum Pub ISBN 1-55591-123-4. PMC 25747993. (Noiz kontsultatua: 2019-12-12).
- ↑ DeMaster, Douglas P.; Stirling, Ian. (1981-05-08). «Ursus maritimus» Mammalian Species (145): 1. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ a b (Ingelesez) Lindqvist, C.; Schuster, S. C.; Sun, Y.; Talbot, S. L.; Qi, J.; Ratan, A.; Tomsho, L. P.; Kasson, L. et al.. (2010-03-16). «Complete mitochondrial genome of a Pleistocene jawbone unveils the origin of polar bear» Proceedings of the National Academy of Sciences 107 (11): 5053–5057. doi:. ISSN 0027-8424. PMID 20194737. PMC PMC2841953. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Waits, Lisette P.; Talbot, Sandra L.; Ward, R. H.; Shields, G. F.. (2008-07-07). «Mitochondrial DNA Phylogeography of the North American Brown Bear and Implications for Conservation» Conservation Biology 12 (2): 408–417. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Marris, Emma. (2007-03). «The species and the specious» Nature 446 (7133): 250–253. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Vermeij, Geerat J.; Motani, Ryosuke. (2018-05). «Land to sea transitions in vertebrates: the dynamics of colonization» Paleobiology 44 (2): 237–250. doi:. ISSN 0094-8373. (Noiz kontsultatua: 2019-12-12).
- ↑ Berta, Annalisa.. (1999). Marine mammals : evolutionary biology. Academic Press ISBN 0-12-093225-3. PMC 42467530. (Noiz kontsultatua: 2019-12-12).
- ↑ a b c d e f (Ingelesez) Schipper, J.; Chanson, J. S.; Chiozza, F.; Cox, N. A.; Hoffmann, M.; Katariya, V.; Lamoreux, J.; Rodrigues, A. S. L. et al.. (2008-10-10). «The Status of the World's Land and Marine Mammals: Diversity, Threat, and Knowledge» Science 322 (5899): 225–230. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2019-12-12).
- ↑ a b c d Riedman, Marianne.. (1990). The pinnipeds : seals, sea lions, and walruses. University of California Press ISBN 0-520-06497-6. PMC 19511610. (Noiz kontsultatua: 2019-12-12).
- ↑ Whitehead, Hal.. (2003). Sperm whales : social evolution in the ocean. University of Chicago Press ISBN 0-226-89517-3. PMC 51242162. (Noiz kontsultatua: 2019-12-12).
- ↑ Dugong : status report and action plans for countries and territories. United Nations Environment Programme [2002] ISBN 92-807-2130-5. PMC 51040880. (Noiz kontsultatua: 2019-12-12).
- ↑ Silverstein, Alvin.. (1995). The sea otter. (Milbrook Press lib. ed. argitaraldia) Millbrook Press ISBN 1-56294-418-5. PMC 30436543. (Noiz kontsultatua: 2019-12-12).
- ↑ Kenyon, Karl W.. (1975). The sea otter in the eastern Pacific Ocean. Dover Publications ISBN 0-486-21346-3. PMC 1504461. (Noiz kontsultatua: 2019-12-12).
- ↑ Stirling, Ian.. (1988). Polar bears. University of Michigan Press ISBN 0-472-10100-5. PMC 17873883. (Noiz kontsultatua: 2019-12-12).
- ↑ Global Business Administration Journal 2 (1) 2018-06-04 doi:. ISSN 2520-9019. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Hochachka, P.; Storey, K.. (1975-02-21). «Metabolic consequences of diving in animals and man» Science 187 (4177): 613–621. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2019-12-12).
- ↑ Cetacean societies : field studies of dolphins and whales. University of Chicago Press 2000 ISBN 0-226-50340-2. PMC 42309843. (Noiz kontsultatua: 2019-12-12).
- ↑ a b Au, Whitlow W. L.. (2000). Hearing by Whales and Dolphins. Springer New York ISBN 978-1-4612-1150-1. PMC 840278009. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Nummela, Sirpa; Thewissen, J.G.M.; Bajpai, Sunil; Hussain, Taseer; Kumar, Kishor. (2007-06). «Sound transmission in archaic and modern whales: Anatomical adaptations for underwater hearing» The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology 290 (6): 716–733. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Reidenberg, Joy S.. (2007-06). «Anatomical adaptations of aquatic mammals» The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology 290 (6): 507–513. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ Klinowska, M.. (1991). Dolphins, porpoises, and whales of the world : the IUCN red data book. IUCN ISBN 2-88032-936-1. PMC 24110680. (Noiz kontsultatua: 2019-12-12).
- ↑ a b Encyclopedia of marine mammals. (2nd ed. argitaraldia) Elsevier/Academic Press 2009 ISBN 978-0-08-091993-5. PMC 316226747. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Sanders, Jon G.; Beichman, Annabel C.; Roman, Joe; Scott, Jarrod J.; Emerson, David; McCarthy, James J.; Girguis, Peter R.. (2015-11). «Baleen whales host a unique gut microbiome with similarities to both carnivores and herbivores» Nature Communications 6 (1): 8285. doi:. ISSN 2041-1723. PMID 26393325. PMC PMC4595633. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Vogl, A. Wayne; Lillie, Margo A.; Piscitelli, Marina A.; Goldbogen, Jeremy A.; Pyenson, Nicholas D.; Shadwick, Robert E.. (2015-05). «Stretchy nerves are an essential component of the extreme feeding mechanism of rorqual whales» Current Biology 25 (9): R360–R361. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ Goldbogen, Jeremy. (2010). «The Ultimate Mouthful: Lunge Feeding in Rorqual Whales» American Scientist 98 (2): 124. doi:. ISSN 0003-0996. (Noiz kontsultatua: 2019-12-12).
- ↑ Marine mammals of eastern North Pacific and Arctic waters. (2nd ed., rev. argitaraldia) Pacific Search Press 1986 ISBN 0-931397-11-1. PMC 13760343. (Noiz kontsultatua: 2019-12-12).
- ↑ VanBlaricom, G. R. (Glenn R.), 1949-. (2001). Sea otters. Voyageur Press ISBN 0-89658-562-X. PMC 46393741. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Mangel, J. C.; Whitty, T.; Medina-Vogel, G.; Alfaro-Shigueto, J.; Cáceres, C.; Godley, B. J.. (2011-04). «Latitudinal variation in diet and patterns of human interaction in the marine otter» Marine Mammal Science 27 (2): E14–E25. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ The encyclopedia of mammals. Barnes & Noble Books 2001 ISBN 0-7607-1969-1. PMC 48048972. (Noiz kontsultatua: 2019-12-12).
- ↑ «Alaska's polar bears trigger lawsuit from industry» Nature 455 (7209): 13–13. 2008-09 doi:. ISSN 0028-0836. (Noiz kontsultatua: 2019-12-12).
- ↑ a b Amstrup, Steven C.; Marcot, Bruce G.; Douglas, David. (2007). «Forecasting the range-wide status of polar bears at selected times in the 21st century» Administrative Report: 1–126. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ Guravich, Dan.. (1993). Polar bear. Chronicle Books ISBN 0-8118-0500-X. PMC 27266879. (Noiz kontsultatua: 2019-12-12).
- ↑ Hemstock, Annie Wendt.. (1999). The polar bear. Capstone High/Low Books ISBN 0-7368-0031-X. PMC 38862448. (Noiz kontsultatua: 2019-12-12).
- ↑ Marsh, H. (Helene), 1945-. (2011). Ecology and conservation of the Sirenia : dugongs and manatees. Cambridge University Press ISBN 978-1-139-16092-6. PMC 783840965. (Noiz kontsultatua: 2019-12-12).
- ↑ Fauna of Australia.. Australian Govt. Pub. Service 1987-<c1989> ISBN 0-644-05483-2. PMC 18523776. (Noiz kontsultatua: 2019-12-12).
- ↑ Estes, J. A.. (1998-10-16). «Killer Whale Predation on Sea Otters Linking Oceanic and Nearshore Ecosystems» Science 282 (5388): 473–476. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ Jokela, Anneli; Arnott, Shelley E.; Beisner, Beatrix E.. (2017-02-07). «Biotic resistance of impact: a native predator (Chaoborus) influences the impact of an invasive predator (Bythotrephes) in temperate lakes» Biological Invasions 19 (5): 1495–1515. doi:. ISSN 1387-3547. (Noiz kontsultatua: 2019-12-12).
- ↑ a b VanBlaricom, G. R. (Glenn R.), 1949-. (2001). Sea otters. Voyageur Press ISBN 0-89658-562-X. PMC 46393741. (Noiz kontsultatua: 2019-12-12).
- ↑ «LETTERS/LETTRES» Canadian Journal of Fisheries and Aquatic Sciences 37 (2): 295–297. 1980-02 doi:. ISSN 0706-652X. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Lunn, Nicholas J.; Servanty, Sabrina; Regehr, Eric V.; Converse, Sarah J.; Richardson, Evan; Stirling, Ian. (2016-07). «Demography of an apex predator at the edge of its range: impacts of changing sea ice on polar bears in Hudson Bay» Ecological Applications 26 (5): 1302–1320. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ a b c Stirling, Ian.. (1988). Polar bears. University of Michigan Press ISBN 0-472-10100-5. PMC 17873883. (Noiz kontsultatua: 2019-12-12).
- ↑ Matkin, Craig O.; Ward Testa, J.; Ellis, Graeme M.; Saulitis, Eva L.. (2013-06-13). «Life history and population dynamics of southern Alaska resident killer whales (Orcinus orca)» Marine Mammal Science 30 (2): 460–479. doi:. ISSN 0824-0469. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Pyle, Peter; Schramm, Mary Jane; Keiper, Carol; Anderson, Scot D.. (1999-04). «PREDATION ON A WHITE SHARK (CARCHARODON CARCHARIAS) BY A KILLER WHALE (ORCINUS ORCA) AND A POSSIBLE CASE OF COMPETITIVE DISPLACEMENT» Marine Mammal Science 15 (2): 563–568. doi:. ISSN 0824-0469. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Visser, Ingrid N.. (2005-01-02). «First Observations of Feeding on Thresher (Alopias vulpinus) and Hammerhead (Sphyrna zygaena) Sharks by Killer Whales (Orcinus orca) Specialising on Elasmobranch Prey» Aquatic Mammals 31 (1): 83–88. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Ford, John K. B.; Reeves, Randall R.. (2008-01). «Fight or flight: antipredator strategies of baleen whales» Mammal Review 38 (1): 50–86. doi:. ISSN 0305-1838. (Noiz kontsultatua: 2019-12-12).
- ↑ Heimlich, Sara.. (2001). Killer whales. (Updated ed. argitaraldia) Voyageur Press ISBN 0-89658-545-X. PMC 46973039. (Noiz kontsultatua: 2019-12-12).
- ↑ Perrin, jean. (1909). «Mouvement brownien et grandeurs moléculaires» Le Radium 6 (12): 353–360. doi:. ISSN 0370-3223. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Springer, A. M.; Estes, J. A.; van Vliet, G. B.; Williams, T. M.; Doak, D. F.; Danner, E. M.; Forney, K. A.; Pfister, B.. (2003-10-14). «Sequential megafaunal collapse in the North Pacific Ocean: An ongoing legacy of industrial whaling?» Proceedings of the National Academy of Sciences 100 (21): 12223–12228. doi:. ISSN 0027-8424. PMID 14526101. PMC PMC218740. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) DeMaster, Douglas P.; Trites, Andrew W.; Clapham, Phillip; Mizroch, Sally; Wade, Paul; Small, Robert J.; Hoef, Jay Ver. (2006-02). «The sequential megafaunal collapse hypothesis: Testing with existing data» Progress in Oceanography 68 (2-4): 329–342. doi:. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Estes, J.A.; Doak, D.F.; Springer, A.M.; Williams, T.M.. (2009-06-27). «Causes and consequences of marine mammal population declines in southwest Alaska: a food-web perspective» Philosophical Transactions of the Royal Society B: Biological Sciences 364 (1524): 1647–1658. doi:. ISSN 0962-8436. (Noiz kontsultatua: 2019-12-12).
- ↑ a b (Ingelesez) Roman, Joe; McCarthy, James J.. (2010-10-11). Roopnarine, Peter ed. «The Whale Pump: Marine Mammals Enhance Primary Productivity in a Coastal Basin» PLoS ONE 5 (10): e13255. doi:. ISSN 1932-6203. PMID 20949007. PMC PMC2952594. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Roman, Joe; Estes, James A; Morissette, Lyne; Smith, Craig; Costa, Daniel; McCarthy, James; Nation, Jb; Nicol, Stephen et al.. (2014-09). «Whales as marine ecosystem engineers» Frontiers in Ecology and the Environment 12 (7): 377–385. doi:. ISSN 1540-9295. (Noiz kontsultatua: 2019-12-12).
- ↑ Smith, Craig; Snelgrove, Paul. (2002-08-29). «A Riot of Species in An Environmental Calm» Oceanography and Marine Biology - An Annual Review (CRC Press): 311–342. ISBN 978-0-415-25462-5. (Noiz kontsultatua: 2019-12-12).
- ↑ (Ingelesez) Fujiwara, Yoshihiro; Kawato, Masaru; Yamamoto, Tomoko; Yamanaka, Toshiro; Sato-Okoshi, Waka; Noda, Chikayo; Tsuchida, Shinji; Komai, Tomoyuki et al.. (2007-03). «Three-year investigations into sperm whale-fall ecosystems in Japan» Marine Ecology 28 (1): 219–232. doi:. ISSN 0173-9565. (Noiz kontsultatua: 2019-12-12).
Kanpo estekak
aldatu Datuak: Q382441
Datuak: Q382441 Multimedia: Marine mammals / Q382441
Multimedia: Marine mammals / Q382441